Autor: Milica Krčmar
Mentor: prof. dr Biserka Vujičić
Fitinska kiselina
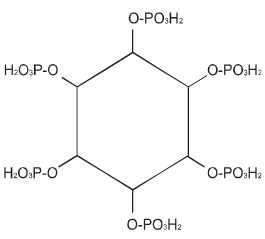
Fitinska kiselina, mioinozitol heksafosfat (Slika 4), odnosno fitin, tj. Ca/Mg so fitinske kiseline, nalazi se u mnogim žitaricama i leguminozama, kao skladišni oblik fosfora i energije gde ova kiselina čini 1% SM ili više i 50-80% fosfora iz biljke je u obliku fitinske kiselina. Fitinska kiselina je prisutna u eukariotskim vrstama i igra značajnu ulogu u metabolizmu eukariota, uključujući i to što je skladišni oblik za fosfor i minerale i što tako i utiče na razvoj i napredak samog semena (Guttieri et al., 2004). Kompleksi fitinske kiseline i mnogih metala, gde se ova kiselina ponaša kao helator nekih jona (cinka, gvožđa, kalcijuma, bakra i drugih) izrazito su stabilni u širokom rasponu pH-vrednosti. Zapravo ovi helati, su depoziti značajni tokom razvoja biljke. Jedan molekul fitinske kiseline može da veže do šest dvovalentnih katjona, uz paralelnu mogućnost intermolekulskog vezivanja dva ili više molekula fitinske kiseline uz učešće katjona, što objašnjava pojavu simptoma nedostatka nekih minerala (najčešće cinka, gvožđa i mangana) u organizmu životinja, naročito svinja i živine, u slučaju kada su obroci za životinje sastavljani isključivo na bazi biljnih komponenti. Pored izrazite tendencije ka formiranju kompleksnih jedinjenja sa metalnim jonima, fitinska kiselina pokazuje sklonost i ka formiranju kompleksnih jedinjenja sa nekim makromolekulima, npr. proteinima, čime takođe utiče na sniženje iskorišćenja ovih nutritienata. Osobina fitinske kiseline da vezuje nutritivno važne komponente obroka u relativno slabo rastvorne, odnosno za životinje neiskoristive komplekse, svrstava ovu komponentu mnogih biljnih hraniva u antinutritiente. Fitinska kiselina je poznata kao antinutrijent zbog tih mogućnosti da kompleksira mikro i makro elemente. Pri neutralnoj pH vrednosti fitinska kiselina je negativno naelektrisna. Iz tog razloga ona privlači više pozitivno naelektrisane grupe kao što su katjoni i proteini. Katjoni se mogu vezati na jednu ili dve fosfatne grupe molekula fitinske kiseline, dok interakcija fitinske kiseline sa proteinima zavisi od pH vrednosti. U visokim koncentracijama može čak da redukuje bio-funkcije minerala i proteina i može kompleksirati sa jon-kofaktorima enzima, koji su neophodni za optimalnu enzimsku aktivnost. U svakom slučaju, u malim koncentracijama, fitinska kiselina može imati neke povoljne nutritivne efekte, uključujući redukciju nivoa usvojivosti skroba i snižavanje nivoa glukoze u krvi, kontrola zubnog karijesa, suzbijanje kancera kolona (bakterije u kolonu produkuju slobodne kiseonikove radikale u značajnim količinama). Dijetetska fitinska kiselina može suzbijati oštećenje od strane kiseonikovih slobodnih radikala u intestinalnom epitelu i susednim ćelijama i obezbeđivati kiseonik neophodan za crvena krvna zrnca. Fitinska kiselina predstavlja prirodni čistač (izmenjivač) za otpadnu vodu, što se koristi i u industrijskim razmerama, za prečišćavanje vode od hroma i bakra u proizvodnji ploča za prečišćavanje (vezuje skoro tri puta više teških metala nego slična količina petroleuma bazne sulfonovane stiren-divinilbenzen smole) (Veličković i sar., 1999).
{kind=link}
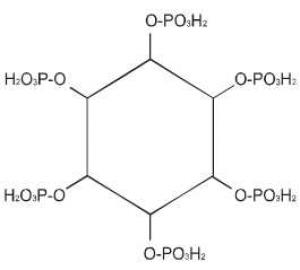
Slika 4. Struktura mioinozitol heksafosfata (fitinske kiseline)
Većina žitarica i uljarica sadrži 1-2% fitata, odnosno 60-90% od ukupnog fosfora čini fitinski fosfor u tim biljnim kulturama. Fitinski fosfor je neiskoristiv za brojlerske piliće, kao uostalom, i za druge vrste monogastičnih životinja, s obzirom da oni ne poseduju fitaze, enzime neophodne za prevođenje fitina u niže fosfatne estre, odnosno mioinozitol. Dodavanje fitaze u obroke za životinje, veoma aktuelno u svetu, kako bi se iskoristio fitinski fosfor i izbegli troškovi vezani za upotrebu dikalcijumfosfata ili nekog drugog mineralnog hraniva kao izvora fosfora za životinje. Ovo zahteva poznavanje sadržaja fitata kako u hranivima, tako i u potpunim krmnim smešama. Najnovija otkrića vezana za antioksidativnu prirodu fitinske kiseline i njen potencijal u lečenju kardiovaskularnih i kancerogenih oboljenja doprinose saznanju o neophodnosti uvođenja brze i jednostavne medote za određivanje sadržaja fitinske kiseline.
Sadržaj fosfora i fitata i njihova distribucija u pasulju kao i u žitaricama je različita u zavisnosti od sorte, a pored toga veliki uticaj ima i količina dodavanog fosfora u đubrivu. Sadržaj fosfora kreće se od 0,4-0,6%, a slični rezultati nađeni su i u literaturi. Sadržaj fitata kreće se od 0,7-1,48%, a u 20 različitih sorti soje kreće se od 18,8-27,7 g/kg. Sadržaj fitata u 14 različitih sorti soje uzgajane u Brazilu kretao se od 0,56-1,20%. U drugim dvema sortama brazilskog pasulja pronađen je sadržaj fitinske kiseline od 1,30 do 1,67%. Drugi autori pronalaze varijacije u sadržaju fitata u različitim usevima kao što su kukuruz i pšenica (Coelho et al., 2002).
Generalno, sadržaj fitata u leguminozama i žitaricama veći je što je veća količina dodatog fosfora u đubrivu. Pronađeno je, u okviru više studija, da se sadržaj fitata u pasulju povećava za 10% sa većim dozama neorganskog fosfora u đubrivu. Bitna činjenica tokom uzgajanja je redukcija fitata u žitaricama bez redukovanja sadržaja fosfora. Iz tog razloga je bitno određivanje frakcija fosfora u formi fitata. Ovaj parametar pruža bitne informacije o tome kada je manje ili više fosfora, u zrnu, prešlo u formu fitata ili neki drugi oblik fosfora i to može pružiti zaključak o regulaciji fitata i njihovoj sintezi. Ispitivanjem 50 sorti pasulja, ustanovljeno je da je sadržaj fitata u semenu bio između 51 i 82% (Coelho et al., 2002).
Neorganski fosfor (P) je drugi važan oblik fosfora u žitaricama. Iako je prisutan u manjim koncentracijama u tom obliku i predstavlja malu frakciju ukupnog fosfora. Visok nivo neorganskog fosfora je označen kao nepoželjan sa nutritivnog aspekta. Zaključci izvedeni iz nekoliko studija o metabolizmu fitinske kiseline bili su da je u pitanju vrlo osetljiva sinteza na nivou ćelije, čija je regulacija slabo poznata, a osnova za sintezu je neorganski fosfor. Sinteza fitinske kiseline može biti regulisana na dva načina. Prvo, fotoasimilacijom fosfora u određenim iznosima i translokacijom fosfora u biljci. Drugi, deljenjem fosfora kao supstrata između različitih depoa i konkuretnih metaboličkih puteva u biljci. Uobičajeno bliska veza između neorganskog P i fitinske kiseline tokom razvoja žitarica i kod odrasle biljke je pronađena, iako je u slučaju genetički modifikovanog kukuruza, redukovan nivo fitinske kiseline, što je pratilo porast neorganskog P. Povećana akumulacija fitinske kiseline može se regulisati od strane fosfoinozitol-kinaze. Ovaj enzim je lociran u važnim tačkama u okviru putanje koja vodi do fitinske kiseline, a neorganski fosfor može direktno da poveća aktivnost ovog enzima koji opet povećava nivo 1L-mio-inozitol-1-P i ATP-a (Coelho et al., 2002).
Gajenje žitarica sa redukovanim nivoom fitinske kiseline može rezultirati neobjašnjivim efektima kao što su redukcija sadržaja P, proteina, mineralnih materija.
U cilju redukcije fitinske kiseline pribegava se i takvim merama kao što su radijacija (zračenje). Proces zračenja hrane se pokazao kao uspešan u smanjenju sadržaja antinutritivnih materija, što obezbeđuje sigurnost i kvalitet hrane kao i zadovoljstvo potrošača. Ova tehnologija je uspešno komercijalizovana od strane vlada 40 zemalja (Bhat et al., 2007) .
Antinutrijenti, posebno fitinska kiselina, smanjuju biološku vrednost dvovalentnih katjona vezujući ih u formu nerastvorljivih fitata. Imperativ je smanjiti nivo fitata, kao i drugih antinutrijenata, odgovarajućim procesnim metodama i osigurati kvalitet namirnice. Sadržaj antinutritivnih materija moguće je smanjiti: natapanjem, sušenjem, kuvanjem, fermentacijom, klijanjem. Poznato je da ovi procesi snižavaju nivo antinutrijenata, naročito fitinske kiseline. Najefektivniji je tretman fermentacije i klijanje, ali je upotreba ovih metoda ograničena, jer zahteva veće radno operećenje zaposlenih što utiče i na cenu ili zbog ograničenja u senzorskom smislu. Potapanje je vid kućne tehnologije, vrlo često u upotrebi u cilju pripremanja obroka. Potapanja ima svoje prednosti jer olakšava kasnije pranje, olakšava i industrijsku proizvodnju i bubrenje semena. Ranije studije (Duhan et al., 2002; Sanberg & Svanberg, 1991) pokazuju da duže natapanje pre fermentacije redukuje sadržaj fitata i menja indeks bioaktivnosti minerala. Smanjenje sadržaja fitata u zavisnosti od sorte i vremena natapanja prikazan je u Tabeli 10. Rezultati pokazuju da različito vreme potapanja može sniziti nivo antinutritijenata ispod dozvoljenih vrednosti (ElMaki et al., 2007).
Duži period potapanja prouzrokuje veći gubitak fitinske kiseline. Gubitak fitinske kiseline u belom pasulju tokom natapanja u vodi posledica je postojanja koncentracionog gradijenta tj. razlike u koncentraciji jona fitinske kiseline u vodi za natapanje i u samom pasulju. Slični rezultati redukcije fitinske kiseline usled potapanja u vodi su od ranije poznati. Uobičajeno kuvanje, potopljenog ili nepotopljenog pasulja pokazuje značajne razlike u sadržaju fitinske kiseline što je prikazano u Tabeli 11. Redukcija fitinske kiseline je primećena i u pasulju koji je kuvan, a da prethodno nije bio potapan u vodu ali su ti gubici manji nego kod pasulja koji je prethodno bio potapan, a potom kuvan.
Prema tome, razlike u gubicima fitinske kiseline tokom kuvanja mogu se objasniti na bazi aktivnosti fitata na temperaturama 40-55°C koje mogu da razgrade inozitol heksafosfat do pentafosfata ili nižih molekulskih formi. Kumar (1978) uočio je da je sniženje sadržaja fitinske kiseline tokom kuvanja posledica formiranja nerastvornih kompleksa fitata s drugim koponentama a samim tim se smanjuje i sadržaj slobodnih fitata. Sadržaj polifenola u različitim sirovim sortama varira od 219 do 676 mg/100 g SM. Za sve sorte, sadržaj polifenola prati promene sadržaja fitinske kiseline posle potapanja (Tabela 13) ili potapanja i kuvanja pasulja (Tabela 14) (ElMaki, 2007).
Tabela 13. Uticaj natapanja na sadržaj fitinske kiseline i polifenolnih jedinjenja (mg/100g SM) u različitim sortama pasulja (ElMaki et al., 2007).
tabela……………………….
Tabela 14. Efekti potapanja i/ili kuvanja na sadržaj fitinske kiseline i polifenola (mg/100 g) u različitim sortama pasulja SM (ElMaki et al.,2007)
tabela……………………………
Mala nutritivna vrednost leguminoza posledica je prisusutva nekih antinutritivnih materija. Tanini onemogućavaju usvajanje proteina, a fitinska kiselina redukuje bioaktivnost nekih esencijalnih mineralnih materija. Može se zaključti, na osnovu ranijih radova, da različite termičke metode obezbeđuju nutricioni kvalitet leguminoza u širem pogledu.
Kataria (1989) izveštava da je kuvanje pod pritiskom efektivnije od običnog kuvanja u redukciji antinutrijenata u crnom i mung pasulju. Saznanja Kadama su (1987) da procesi ključanja i obrade u autoklavima značajno redukuju antinutritivne komponente i poboljšavaju kvalitet proteina. Mbofung (1999) kaže da iskoristljivost skroba direktno posledica kuvanja na pari kao metode termičke obrade. Pored toga što se kuvanjem smanjuje količina antinutritivnih materija, smanjuje se i količina prisutnih mineralnih materija, što se može okarakterisati kao nepoželjna pojava. Kuvanje i klijanje dovodi i do smanjenja masnoća, frakcija ugljenih hidrata, antinutritivnih faktora i pepela. Svi procesi, za smanjenje antinutritivnih materija, dovode do snižavanja količine lizina, triptofana, treonina i sulfo-aminokiselina. Ali, s druge strane povećava se sadržaj aromatičnih amino kiselina, leucina, izoleucina i valina. Procesi kuvanja su efektivniji od klijanja u pogledu redukcije tripsin inhibitora, tanina i hemaglutinina, dok je klijanje u odnosu na kuvanje efektivnije u pogledu redukcije fitinske kiseline, stahioze i rafinoze (Mubarak et al., 2005). Za sve leguminoze je karakteristično i uobičajeno jednostavna metoda kuvanja u ključaloj vodi ili pod pritiskom (Rehman et al., 2005).
.
Određivanje fitinske kiseline
Prisustvo fitinske kiseline u mnogim vrstama biljaka, pšenici, kukuruzu, riži, soji dobro je poznata i dokumentovana. Fitinska kiselina je mio-inozitol, fosforilizovana na svih 6 hidroksilinih grupa. Fitinska kiselina je najčešće u obliku Ca/Mg soli.
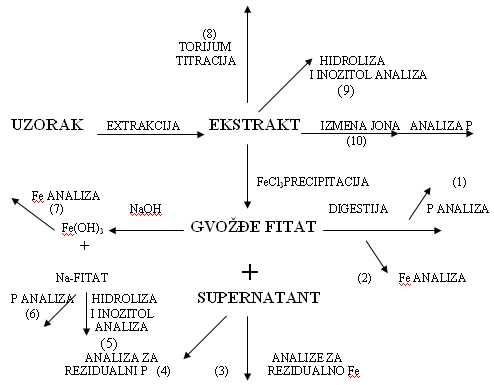
Fitinska kiselina je konstituent žitarica i proizvoda od žitarica. Poznato je da ona umanjuje bioaktivnost cinka i drugih elemenata u tragovima kod ljudi i monogastičnih životinja (Oberlease et al., 1961; O’Dell et al., 1960; Reinhold et al., 1973). Zbog toga je sadržaj i distribucija fitinske kiseline u žitaricama od velikog ineresa, a brza i osetljiva metoda analize fitinske kiseline zahteva upotrebu malih uzoraka. U konvencionalnim metoda fitinska kiselina se taloži kao gvožđe-fitat. Talog se separira, ispere, i sadržaj fosfora odredi nakon hidrolize. Sadržaj fitinske kiseline se izračunava preko sadržaja fosfora. Ovaj metod zahteva mnogo vremena.
Mnogo brži način za određivanje fitinske kiseline je dobijen modifikacijom originalne metode po Young-u (1936). U tom smislu se dobijeni ekstrakt uzorka zagreva sa zakišeljenim rastvorom trovalentnog gvožđa poznate koncentracije. Smanjenje sadržaja gvožđa u supernatantu (određuje se kolorimetrijski sa 2,2′-bipiridinom) je mera za sadržaj fitinske kiseline. Metoda je indirektna jer se vrši kalibracija uz pomoć rastvora fitinske kiseline poznate koncentracije (standardni rastvori). Slična metoda za određivanje fitata u soji opisana je od strane Davies-a (1979) koji je modifikovao metodu koju je prethodno razvio Holt (1955). U ovoj metodi, gvožđe se određuje tiocijanatom ali ovaj kompleks se razara na svetlosti što nam onemogućava dobijanje validnih rezultata u ekstraktima ceralija (Haug et al., 1983).
U ovom radu je kao relevantana metoda izabrana modifikovana metoda Haug, W. & Lantzsch, H-J (1983): Metoda za brzo određivanje fitata u žitaricama i proizvodima od žitarica. U nekim predhodnim određivanjima (Tepić i sar., 2007) utvrđeno je da ova metoda daje najbolje rezultate u okviru naših laboratorijskih uslova. Metoda se zasniva na indirektnom kolorimetrijskom određivanju fitinskog fosfora.
Fitinska kiselina se taloži dodavanjem feri-amonijumsulfata poznate koncentracije gvožđa. Deo gvožđa se troši na stvaranje feri-fitata a preostali deo gvožđa u supernatantu se određuje spektrofotometrijski. Metoda je indirektna jer se određivanje vrši u odnosu na kalibracionu krivu formiranu uz pomoć standardnih rastvora natrijumove soli fitinske kiseline.
U upotrebi je nekoliko različitih metoda za određivanje fitinske kiseline a, koje ne zahtevaju sofisticiranu opremu. Ove metode su razvijene u cilju određivanja sadržaja fitinske kiseline u različitim materijalima kao što su: biljne ćelije, hrana i zemljišta. Određivanje fitata izmenom jona je odobrena oficijalna AOAC metoda. Ograničena je dugim vremenskim određivanjem što je nepodesno za rutinsko određivanje u slučaju većeg broja uzoraka (AOAC, 1995). U proteklim godinama, razvijeno je nekoliko HPLC metoda za određivanje fitata. Upotrebom gradijenta izmenjivača jona u okviru HPLC, upotrebom odgovarajućeg detekcionog sistema (detekcija nakon razdvajanja) u kolonama, obezbeđuje separaciju mio-inozitol fosfata uključujući fitinsku kislinu. Heksa-, penta-, tetra-, tri-, i difosfati članovi ove homologe serije jasno su određeni u roku od 30 minuta. Ovaj metod bi trebao da olakša analizu i kvantitativno određivanje fitinske kiseline i drugih inozitol fosfata u biljkama, hrani i zemljištima. Sve razvijene HPLC metode zahtevaju upotrebu dodatnog detekcionog sistema u kolonama sastavljenog od pumpe sa reagensom i ankera sa namotanom žicom i sa mešanjem za razvijanje boje. Jednom instalisan i kalibrisan HPLC sistem je podesan za određivanje većeg broja uzoraka.
Makro metoda određivanja supernatanta koju su razvili Erdman i Thompson (1982) za određivanje fitinske kiseline u soji i proizvodima od soje, prerađena je u mikrometodu. Mikro metoda je razvijena u cilju bržeg i lakšeg određivanja fitinske kiseline. Prednost mikro u odnosu na makro metodu je smanjivanje količine uzorka za analizu što onda zahteva i manje količine reagenasa, a smanjuje se i vreme određivanja. Zamenjujući fazu dugog filtriranja sa kratkotrajnim centrifugiranjem vreme analize je skraćeno sa 14 na 8 h. Fitinska kiselina je određivana u uzorcima soje koresteći obe metode i obrađeni podaci su pokazali visoko slaganje. Tokom istraživanja utvrđeno je da je mikro metoda bila uspešnija od makro metode i podesna za određivanja u izolatima soje (Veličković i sar., 1999).
U cilju da se na adekvatan način utvrdi veza između fitinske kiseline i mineralnih materija u soji, mora se biti u sposobnosti odrediti jednu od te dve komponenete. Nekoliko različitih metoda može se naći u literaturi (Tabela 15). Ove metode su razvijene da bi se odredila količina fitinske kiseline u najrazličitijim materijalima (Thompson et al., 1982).
Mnoge metode su zasnovane na precipitaciji gvožđe-fitata sa gvožđe-hloridom. Metode pokazuju da talog može biti direktno rastvoren i analiziran na fosfate (1) ili na gvožđe (2). Alternativno, natrijum-hidroksid može biti korišćen za konverziju precipitata u natrijum-fitat i gvožđe-hidroksid. Zgodno je odrediti gvožđe nakon dodatka gvožđe-hidroksida u kiselinu (7). Određivanje rezidualnog gvožđa u supernantantu (3) je takođe brza metoda. Određivanje rezidualnog fosfora u supernatantu nije kao metoda pominjana u literaturi (druga grupa istraživača, po Tangkongchitr et al., 1981 uvrštava ovu metodu kao deo određivanja ukupnog fosfora i odnosa sa fitinskom kiselinom tokom pečenja hleba).
tabela……………………….
Jonska izmena separirane fitinske kiseline iz ekstrakta (10) je metoda koja je izvođena od strane Herald i Oberlase (1977) na pšeničnim mekinjama. Metoda je ograničena dugim vremenom određivanja što je nepovoljno kod rutinskih određivanja većeg broja uzoraka. Određivanje fitinske kiseline je pod velikim uticajem okruženja tj. vrste namirnice u kojoj se određuje. To je posledica toga što se okruženja u kojima je nađena fitinska kiselina, a u različitim materijalima ne slažu. Fitati pronađeni u različitom okruženju minerala i proteina neće se ponašati precizno isto pri istim upotrebljenim metodama.
{kind=link}
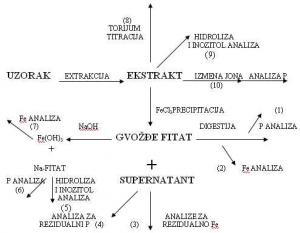
Slika 5. Mogući načini određivanja fitinske kiseline (Thompson & Erdman, 1982)
.
Fitinska kiselina u prosu
Antinutrijenti (fitinska kiselina i polifenoli) prisutni su u nezanemarljivim iznosima i utiču na ograničenu usvojivost skroba i proteina, i na bioaktivnost minerala. Inhibiraju proteolitičke i amilolitičke enzime.
Tabela 16. Povezanost fitinske kiseline i ukupnog fosfora u prosu (Abdalla et al., 1997)
tabela…………………………..
Fitinska kiselina u prosu određena je metodom koju su opisali Wheeler & Ferrel (1971). Rezultati su izraženi na suvu materiju. Kao što se vidi u Tabeli 13, sadržaj fitinske kiseline se kreće od 354 do 769 mg/g. Varijacije u sadržaju posledica su različitih genotipova i različitih uticaja iz okoline (Abdalla et al., 1998).
.
Fitinska kiselina u pšenici
Analize sadržaja frakcija fosfora u semenu pšenice:
Analitički protokol za određivanje kvantitativnog hemijskog sastava je poznat i opisan (Larson et al., 2000; Raboy et al., 2000; Dorsch et al., 2003). Ukupan fosfor određen je kolorimetrijski (Chen et al., 1956) iz uzorka u formi pepela, fitinska kiselina je određena precipitacijom sa gvožđem a neorganski fosfor kolorometrijski ekstrakcijom sa 0,25 magnezijum-hloridom u 12,5% 2,4,6-trihloranizolom.
Anjonska izmena u okviru HPLC metode, sa detekcijom nakon razdvajanja je opisana od strane (Larson et al., 2000; Raboy et al., 2000; Dorsch et al., 2003). Vršena je ekstrakcija sa HCl, supernatant razblažen sa bidestilovanom vodom i profiltriran. Alikvot je frakcionisan u Dionex IonPac AS7 jonskom izmenjivaču opremljenim sa Dionex IonPac AG7 kolonom. Sadržaj fosfora izračunat na osnovu kalibracione krive formirane sa standardnim rastvorima (Guttieri et al., 2004).
Tabela 17. Koncentracija fosfora u prolećnoj pšenici, samlevenim frakcijama, proizvedenoj u Tetoniji 2001. (Guttieri et al., 2004)
tabela………………………….
.
Fitinska kiselina u bobu
Fitinska kiselina je ekstrahovana iz uzoraka semena sa 2,4% HCl. Dobijena smeša je centrifugirana pri ubrzanju 17,300 g (Sorvalle RC-5B) 30 minuta pri 15ºC i prikupljeni su supernatanti. Stakleni cilindri, Econo-colone 0,7 x 15 cm (Bio Rad laboratorija), isprani su sa 0,5 g AG 1 x 4 anjonskim izmenjivačem kako bi se olakšala purifikacija fitata. Postupak određivanja ukupnog sadržaja fitinske kiseline je zasnovan na reakciji gvožđe(III)-hlorida sa sumpor-salicilnom kiselinom (Alonso et al., 2000).
Tabela 18. Fitinska kiselina, tanini i polifenoli (g/kg SM) u bobu (Vicia faba L.) i pasulju (Phaseolus vulgaris L.) (Alonso et al., 2000)
tabela…………………….
.
Fitinska kiselina u lanenom semenu
Sadržaj fitinske kiseline u lanenom semenu (Linum usitatissimum) određen je prema metodi Wheeler-a i Ferrel-a (1971). Rezultati ukupnog istraživanja prikazani su u tabeli 19. Eliminišući/redukujući tanine i fitinsku kiselinu, adekvatno vođenom fermentacijom postiže se bolja usvojivost proteina i masnoća (Mukhopadhyay, 2005).
Tabela 19. Prosečni sastav semena lana (u % izraženo na suvu materiju) (Mukhopadhyay, 2005).
tabela………………………….
.
Fitinska kiselina u soji
Fitinska kiselina prisutna u semenu soje (Glycine max L. Merill) je prilično termostabilna komponenta koja formira helate sa jonima Ca2+, Mg2+, Zn2+, Fe3+, čime oni postaju manje dostupni (Dai et al., 2007).
Tabela 20. Sadržaj proteina, masnih kiselina, ulja i fitinske kiseline u sedam različitih genotipova soje (Kumar et al., 2006)
tabela……………………..
U ispitivanjima su korištena sveža semena soje sa četri lokacije, sušena na 60°C, usitnjena i prosejana kroz otvore prečnika 0,5 mm, a potom određena fitinska kiselina standardnom metodom Wheeler & Ferrel (1971). Sadržaj fitata je izračunat iz koncentracije gvožđa iz dodatog gvožđe-hlorida, uzimajući u obzir činjenicu da je odnos Fe:P=1:1,5 i da je konstantan. Dodatkom gvožđe-hlorida vrši se taloženje fitata u formi gvožđe-fitata. Rezultati su prikazani u Tabeli 20.
.
Fitinska kiselina u sočivu
Nutritivni kvalitet i sastav su bitni za potrošače jer se sočivo koristi i za ljudsku i za životinjsku ishranu. Nivo fitinske kiseline kreće se od 0,30% do 1,20% sa vrednošću odnosa 0,91% (tabela 21). Različite vrste pokazuju različit sadržaj fitinske kiseline. Fitinska kiselina određena je kolorimetrijski na 500 nm koristeći spektrofotometar (Wang et al., 2006).
Tabela 21. Šećeri, fitinska kiselina, tanini i tripsin inhibitor (TIA) u sočivu (Wang et al., 2006)
tabela……..
Spisak korišćene literature možete naći u Literatura – Hemija hrane.