THE HIGH-MOLECULAR WEIGHT GLUTENIN SUBUNIT (HMW GS) COMPOSITIONS IN CURRENT WHEAT (Tr. aestivum L.) VARIETIES IN BiH
Originalan naučni rad – Original scientific paper
Rezime
Za jedinstveno viskozno-elastično svojstvo tijesta zamješenog od brašna heksaploidne pšenice (Tr. aestivum L) u prvom redu su odgovorni skladišni proteini. Ovo svojstvo je presudno u procesu proizvodnje dizanih pekarskih proizvoda.
Skladišni proteini pšenice sastoje se od gluteninskih subjedinica velike molekulske težine (HMW GS) oko 10%, gluteninskih subjedinica male molekulske težine (LMW GS) oko 40% i oko 50% glijadina. Ustanovljena je izražena alelna varijacija i glijadina i gluteninskih subjedinica, što povećava različitosti između kompozicija skladišnih proteina među genotipovima. Ova varijacija uzrokuje razlike u kvalitetu proteina sorti pšenice.
HMW gluteninske subjedinice čine tek mali udio ukupnih skladišnih proteina, ali ispoljavaju presudan uticaj na elasticitet glutenina i kvalitet pšeničnog hljeba.
U ovom radu je determinisana kompozicija HMW gluteninskih subjedinica nekih aktuelnih sorti pšenice u Bosni i Hercegovini, korištenjem metode elektroforeze na poliakrilamidnom gelu u prisustvu SDS, i prezentirani dobijeni rezultati.
Summary
The storage proteins of hexaploid wheat (Tr. aestivum L.) are primarily responsible for the unique cohesive-elastic properties of dough made from wheat flours. These properties are essential in processing of leavened bread.
The storage proteins of wheat contain about 10% HMW glutenin subunits, about 40% LMW glutenin subunits and about 50% gliadins. Extensive allelic variation in both gliadins and glutenin subunits are detected, giving rise to different composition of storage proteins between genotypes. This variation causes differences in protein quality among varieties.
The HMW subunits of glutenin constitute only a small proportion of the storage proteins, but they exert pronounced effects on the glutenin elasticity and the bread making quality of the wheat flour.
The high molecular weight glutenin subunits (HMW GS) composition of some current wheat varieties in Bosnia and Herzegovina was determined by polyacrylamide gel electrophoresis in the presence of SDS, and presented in this paper.
Key words: wheat, HMW glutenin subunits, SDS-PAGE
Skraćenice/Abbreviations: SDS: Natrijum dodecil sulfat/Sodium dodecyl sulphate; HMW-GS: Subjedinice glutenina velike molekulske težine/High molecular weight glutenin subunits; SDS-PAGE: Elektroforeza na poliakrilamidnom gelu za određivanje subjedinica glutenina velike molekulske težine/Sodium dodecyl sulphate polyacrilamide gel electrophoresis
Uvod
Danas je već sasvim jasno da su, generalno, za proizvodnju hljeba i tehnologiju brašna najvažniji faktori koji karakteriziraju dobru sirovinu kvantitet, i posebno, kvalitet proteina glutena. Dobra sirovina podrazumijeva i stabilan kvalitet brašna. Osigurati stabilan kvalitet brašna osnovni je cilj proizvođača, ali ujedno i veliki izazov, pošto kompoziciju proteina glutena, odnosno njegov kvalitet, uvjetuju genetske predispozicije i uslovi gajenja. Stoga, nije čudo što se veliki broj naučnih istraživanja fokusira upravo na ispitivanje kvaliteta proteina glutena u relaciji sa pecivim osobinama, odnosno, iznalaženju načina da se omogući izvjesno predviđanje kvaliteta hljeba na osnovu kvaliteta glutena.
Odabir kvalitetnih sorti pšenice trebalo bi da bude imperativ za domaće proizvođače pšenice, kako bi se povećala proizvodnja u Bosni i Hercegovini i osigurala „čista“ i kvalitetna sirovina, što je ujedno i preporuka ovog istraživanja.
Ustanovljena veza između kompozicije HMW GS i kvaliteta pekarskih proizvoda može poslužiti prilikom uzgoja, i u selekciji, koristeći metod SDS-PAGE. Ova tehnika elektroforeze je relativno brza i može se provoditi na pojedinačnim zrnima i u ranijim stadijima uzgojnog programa.
Kompozicija HMW GS je značajan pokazatelj kvaliteta glutena, međutim ova vrsta ispitivanja kod nas je nedovoljno ili nikako provođena. Stoga je i cilj rada bio karakterizacija HMW GS u aktualnim sortama pšenice koje su najčešće prisutne u Bosni i Hercegovini.
Pregled literature
Proteini su različito raspoređeni u dijelovima pšeničnog zrna, međutim funkcionalnu ulogu imaju samo skladišni proteini, odnosno proteini endosperma.
Glutenini su nerastvorljivi u alkoholima i sastoje se od proteinskih subjedinica prisutnih u polimerima stabiliziranih među-lančanim disulfidnim vezama. Redukcija ovih veza ima za posljedicu nastajanje subjedinica koje su rastvorljive u smjesi voda/alkohol i klasificiraju se kao dva tipa:
1. HMW (High molecular weight) gluteninske subjedinice
2. LMW (Low molecular weight) gluteninske subjedinice (S h e w r y et al, 1997).
Svaki tip proteina ima molekulsku masu, rastvorljivost, agregaciju i karakteristike djelimično različite od proteina iz drugih grupa. Svaki polipeptidni tip se, takođe, sastoji od velikog broja blisko vezanih proteina, koji nastaju preko duplikacije gena i mutacijom (W r i g l e y i B i e t z, 1988).
Viskozno-elastični karakter zavisi i od polimernih proteina (glutenini), koji doprinose elastičnosti tijesta, i od monomernih proteina (glijadini), koji doprinose rastegljivosti (E l i a s s o n, 1990). Ravnoteža između elastičnosti i rastegljivosti, preko genetskih predispozicija, može jako varirati među sortama, i ima glavni uticaj na tip gotovih proizvoda koji se dobijaju od pšeničnog brašna (P a y n e, 1987).
Hljebna pšenica (Triticum aestivum) je heksaploidna vrsta, koja sadrži tri različita, ali povezana genoma, od sedam hromozomskih parova, nazvanih A, B i D. Svaki od 21 para hromozoma označava se brojem hromozoma od 1 do 7, prateći raspored genoma (A, B ili D). Geni koji kontroliraju HMW subjedinice glutenina pojavljuju se na dugim krakovima hromozoma 1A, 1B i 1D. Geni koji kontroliraju LMW subjedinice glutenina, ω-glijadine i γ-glijadine pojavljuju se na kratkim krakovima istog seta hromozoma, a geni za kontrolu α- i β-glijadina pojavljuju se na kratkim krakovima 6A, 6B i 6D hromozoma (P a y n e, 1987). Geni za γ- i ω-glijadine u grupi 1 kratkog kraka su blisko vezani za gene koji kodiraju LMW gluteninske subjedinice. Svi ovi lokusi izloženi su alelnoj varijaciji (M a c R i t c h i e, 1999).
Lokusi koji sadrže gene koji kontroliraju HMW gluteninske jedinice nazivaju se zajedničkim imenom Glu-1, a posebno se označavaju kao Glu-A1, Glu –B1, i Glu-D1, i odnose se na hromozome 1A, 1B, i 1D (P a y n e, 1987).
Metod za identifikaciju HMW gluteninskih subjedinica opisali su autori Payne i Lawrence (1983). Payneet al (1985) predstavili su dvodimenzionalno frakcionisanje proteina endosperma sa biohemijskom i genetskom studijom, a Lafiandra i Kasarda (1985) opisali su jedno i dvodimenzionalnu elektroforezu u poliakrilnom gelu za separaciju pšeničnih proteina na pojedinačnom gelu.
Na bazi elektroforetske mobilnosti i izoelektričnog fokusiranja, subjedinice kodirane od strane pojedinačnih lokusa dalje se dijele na „x“ i „y“ tipove, a kod subjedinica kodiranih od strane 1A postoji samo jedan tip, x tip (P a y n e, 1987).
1A y-subjedinice nikad nisu nađene kod heksaploidne pšenice, ali su nađene kod nekih divljih diploidnih vrsta (U h l e n, 1989). Neizražena 1A x-subjedinica Glu-A1 alela označava se kao nul alel, i predstavlja primjer nefunkcionalnih gena (pseudogena) u alelima skladišnih proteina (P a y n e, 1987).
Tipovi subjedinica 1Ax, 1Bx, i 1Dx su više slični jedni drugima, u odnosu na 1By i 1Dy subjedinice. Druga razlika kod tipova subjedinica je ta da subjedinice tipa x sadrže za oko pola manju količinu cisteina, u odnosu na subjedinice tipa y (Payne, 1987).
Teorijski, heksaploidna hljebna pšenica bi mogla sadržavati šest različitih subjedinica. U stvari, u kultivarima su prisutne samo tri, četiri ili pet subjedinica, što je posljedica mirovanja pojedinih gena. Pojedinačne subjedinice su numerisane po redoslijedu kako im raste mobilnost prema SDS-PAGE. Ovaj polimorfizam omogućava osnovu za korelaciju specifičnih alela u odnosu na različitosti u pogledu kvaliteta hljeba (S h e w r y et al, 1992).
Radovi brojnih autora, posebno Paynea i saradnika, ukazuju da se glavne subjedinice, koje su kodirane od strane A, B i D genoma, mogu rangirati prema kvalitetu. U pogledu 1D i nekih 1B alela, bliska veza između gena za x- i y-tipove subjedinica upućuje da se ocjena za kvalitet, u prvom redu, odnosi na parove subjedinica, prije nego na subjedinice pojedinačno. Subjedinice najboljeg kvaliteta su par alela 1Dx5+1Dy10, kada se porede sa 1Dx2/3/4+1Dy12. Kod 1Ax subjedinica 1 i 2*, postoji neznatno slabiji kvalitet u usporedbi sa nultim alelom (S h e w r y et al, 1992).
Neznatne razlike u aminokiselinskim sekvencama između subjedinica 5+10 i 2+12 sugerišu objašnjenje za značajne razlike u njihovoj sekundarnoj strukturi. Nadalje, nađeno je da Glu-B1 subjedinice 13+16 osiguravaju bolji kvalitet u odnosu na subjedinice 6+8 (P a y n e et al, 1981).
Iako HMW gluteninske jedinice čine tek oko 10% skladišnih proteina u endospermu, ova frakcija ima odlučujući efekat na pecive osobine i kvalitet hljeba. Istraživanja potvrđuju da je 47-60% variranja u pogledu kvaliteta 84 britanske sorte pšenice moguće objasniti sastavom HMW gluteninskih subjedinica (P a y n e et al, 1987a, prema U h l e n 1990). Ispitivanja provedena na španskim sortama pšenice pokazala su slične rezultate (P a y n e et al, 1988).
Osamdesete godine prošlog stoljeća mogu se nazvati „dekadom HMW subjedinica“ zbog velikog interesovanja za ovu, prema kvantitetu minornu, ali, prema važnosti, veoma značajnu grupu protiena glutena. Ovaj interes je podržan prvenstveno zahvaljujući radovima Paynea i saradnika, koji su opisali korelaciju specifičnih alela subjedinica i kvaliteta evropskih pšenica u pogledu proizvodnje hljeba (S h e w r y et al, 1992).
Slična ispitivanja provedena su od strane brojnih autora (Gupta et al, 1991; Lookhart et al, 1993; P e n a et al, 2005);
Nađeno je da je relativna količina različitih HMW gluteninskih subjedinica približno konstantna kod različitih genotipova, te informacija o prisustvu ili odsustvu različitih alela HMW gluteninskih subjedinica, uz sadržaj proteina, može nositi glavnu informaciju o ukupnoj količini HMW gluteninskih subjedinica u genotipovima (U h l e n, 1990). Rezultati pomenutog autora sugerišu da razlike u biohemijskim svojstvima subjedinica mogu predstavljati značajniji faktor kvaliteta nego ukupna količina HMW gluteninskih subjedinica.
Uticaj variranja lokusa gluteninskih jedinica na maksimalni otpor tijesta ispitivali su G u p t a et al (1994) i rangirali analizirane alele kao: Glu-D1 > Glu-B1 > Glu-B3 > Glu-A3 > Glu-D3 = Glu-A1.
Naučno utemeljeno stajalište o vezi između specifičnih HMW gluteninskih subjedinica i pecivih osobina i kvaliteta hljeba danas uveliko koriste selekcioneri, koristeći SDS-PAGE kao test, koji je relativno brz. Uz to, ova elektroforetska tehnika može se izvesti na veoma maloj količini uzorka, a selekcija se može provoditi još u ranoj fazi selekcionog programa.
Veliki interes za determinaciju HMW gluteninskih subjedinica proizilazi iz njihove pogodnosti za analitičko određivanje, pošto se lociraju na vrhu elektroforetskog gela (prema rutinskoj metodi sa rastvorom SDS) i jasno odvajaju od ostalih polipeptidnih lanaca.
Materijal i metode rada
Istraživanja u radu provedena su laboratoriju Agricultural University of Norway, Department of Crop Science, Norveška.
Uzorci pšenice za ispitivanje, osam aktuelnih sorti, predstavljaju sorte koje su bile najviše zastupljene u BiH tokom 2003. i 2004. godine. Lokaliteti sa kojih su sakupljeni uzorci su Tuzla i Banja Luka, sa oglednih polja. Radi se o ozimim sortama pšenice, namijenjenim uglavnom za pekarsku proizvodnju.
Uzorci pšenice su nakon žetve prosušeni na vazduhu, pri ambijentalnoj temperaturi, kako bi se odstranio višak vlage, i uskladišteni u odgovarajućim uslovima.
Kod svih uzoraka klijavost (K a l u đ e r s k i i F i l i p o v i ć, 1998) je iznosila preko 98%, što potvrđuje odgovarajuće zdravstveno stanje i svježinu zrna.
Količina uzoraka pšenice iznosila je oko 10 kg. Uzorci pšenice koji su uzeti za ispitivanje prikazani su u tabeli 1.
Tabela 1.
Wheat samples with locations and growing seasons denoted
| Sorta pšenice Wheat cultivar | Lokalitet Location | Godina uzgoja Growing season | Oznaka u radu Mark in paper | |
| 1. | Marija | Banja Luka | 2003 | Marija BL 03 |
| 2. | Marija | Banja Luka | 2004 | Marija BL 04 |
| 3. | Marija | Tuzla | 2004 | Marija T 04 |
| 4. | Liberta | Banja Luka | 2003 | Liberta BL 03 |
| 5. | Liberta | Banja Luka | 2004 | Liberta BL 04 |
| 6. | Trna | Banja Luka | 2003 | Tina BL 03 |
| 7. | Trna | Tuzla | 2003 | Tina T 03 |
| 8. | Trna | Banja Luka | 2004 | Tina BL 04 |
| 9. | Trna | Tuzla | 2004 | Tina T 04 |
| 10. | Sana | Banja Luka | 2003 | Sana BL 03 |
| 11. | Sana | Banja Luka | 2004 | Sana BL 04 |
| 12. | Renesansa | Banja Luka | 2003 | Renesansa BL 03 |
| 13. | Renesansa | Tuzla | 2003 | Renesansa T 03 |
| 14. | Renesansa | Tuzla | 2004 | Renesansa T 04 |
| 15. | Rusija | Tuzla | 2003 | Rusija T 03 |
| 16. | Rusija | Tuzla | 2004 | Rusija T 04 |
| 17. | Pobeda | Tuzla | 2003 | Pobeda T 03 |
| 18. | Pobeda | Tuzla | 2004 | Pobeda T 04 |
| 19. | Evropa 90 | Tuzla | 2003 | Evropa T 03 |
| 20. | Evropa 90 | Tuzla | 2004 | Evropa T 04 |
Sorte Marija [(ZG 4527/68 x KAVKAZ) x ZG 1971/70]; Liberta [(M-41-1 x DRINA) x ZG 167/86] (sublinija SANE); Tina (SANA x GALA) i Sana [(MURA x CI 14123) x ZG 2413/72] su selekcionisane na Bc Institutu za oplemenjivanje i proizvodnju bilja d.d. Zagreb, a sorte Renesansa (JUGOSLAVIJA x linije NS 55-25); Rusija (SREMICA x NOVOSADSKA RANA 2); Pobeda (SREMICA x BALKAN) i Evropa 90 (TALENT x NOVOSADSKA RANA 2) potiču iz Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad.
Određivanje HMW-GS kompozicije
Kompozicija subjedinica glutenina velike molekulske težine (HMW-GS) određena je pomoću elektroforeze proteinskog ekstrakta na poliakrilamidnom gelu metodom SDS-PAGE (Sodium dodecyl sulphate polyacrylamede gel electrophoresis) po Payne at al. (1980), sa manjim modifikacijama prema Uhlen (1990). Karakterizacija HMW-GS je rađena u paralelnim određivanjima za sve uzorke.
Elektroforeza predstavlja kretanje suspendiranih ili rastvorenih čestica u primijenjenom električnom polju. Mobilnost čestica zavisi od električnog napona i veličine polipeptida.
Elektroforeza omogućava separaciju i vizualizaciju proteina u uzorku, tokom zadatog vremena. Prirodni glutenin je potrebno prethodno reducirati na subjedinice.
Određivanje kompozicije HMW-GS rađeno je na 20 uzoraka pšenice iz BiH. Kod uzoraka pšenice: Marija BL 03; Liberta BL 03; Liberta BL 04; Tina BL 03; Tina T 03; Sana BL 03 i Renesansa BL 03 ponovljeno je određivanje HMW-GS na pojedinačnim zrnima (po 5 zrna), radi provjere dobivenih rezultata.
Uzorci su prethodno pripremljeni mljevenjem na laboratorijskom mlinu za određivanje broja padanja (Falling Number 3100 Hammer Mill; Perten Instruments AB, Sweden).
Količina od 0,5 ml ekstrakcionog pufera je dodata na 20 mg usitnjenog uzorka. Pufer od 0,063 M Tris-HCl (pH = 6,8) sadrži 2% SDS-a, 5% ditiotri-itola, 20% glicerola i 0,001% Pyronina y.
Sloj poliakril-amidnog gela (10%) nanesen je prema metodi po P a y n e at al (1980), sa 4% čvrstog gela na vrhu.
Elektroforeza je provedena na aparaturi BIO RAD Mini Protean II. Odgovara za trake gela veličine 140 x 180 x 1,5 mm.
5μl uzorka nanosi se na gel. Na pojedinačni gel može se nanijeti 15 uzoraka. Elektroforeza se provodi uz 20mA/2 gela u toku prvih 15 minuta. Postupak se nastavlja uz 40mA/2 gela u trajanju od oko jednog sata, odnosno dok ružičasti obojeni trag (Pyronin y) ne iščezne sa dna gela.
Gelovi se boje sa kumaši plavim (Coomassie Brilliant Blue R-250) u trajanju od 24 sata i potom ispiraju u destilovanoj vodi.
Na svaki gel je uz uzorke nanesen proteinski ekstrakt standardne sorte poznate kompozicije HMW-GS, kako bi se očitavanje preciznije obavilo. Kao standard su korištene norveške sorte: Zebra i Mjølner. Identifikacija HMW-GS je provedena po sistemu prema P a y n e at al (1983).
Za digitalno predstavljanje gelova sa uzorcima i standardima korišten je „Gel Pro“ Program (BIO RAD Mini Protean II).
Rezultati i diskusija
Kvalitativna identifikacija HMW gluteninskih subjedinica predstavljala je predmet ovog istraživanja. Kvantitativna analiza ovih subjedinica nije rađena, a razlog za to je jednostavno identificiranje ove proteinske grupe, a relativan odnos različitih HMW gluteninskih subjedinica je relativno konstantan u uzorcima pšenice (Uhlen, 1990). Ovakva odluka je u skladu sa istraživanjima Faergestad et al (2004). Sastav gluteninskih subjedinica velike molekulske težine (HMW-GS) pojedinih uzoraka pšenice predstavljen je u tabeli 2.
Tabela 2.
High molecular weight glutenin subunits (HMW GS)
| Uzorak Sample | Glu-Al | Glu-Bl | Glu-Dl |
| Marija BL 04 | 2* | 7+9 | 5+10 |
| Marija T 04 | 2* | 7+9 | 5+10 |
| Tina BL 04 | 2* | 6+8 | 2+12 |
| Tina T 04 | 2* | 6+8 | 2+12 |
| Sana BL 04 | 2* | 6+8 | 5+10 2+12 |
| Renesansa T 03 | 2* | 7+9 | 5+10 |
| Renesansa T 04 | 2* | 7+9 | 5+10 |
| Rusija BL 03 | 2* | 7+9 | 2+12 |
| Rusija T 03 | 2* | 7+9 | 5+10 |
| Pobeda BL 03 | 2* | 7+9 | 5+10 |
| Pobeda T 04 | 2* | 7+9 | 5-10 |
| Evropa T 03 | 2* | 7+9 | 2+12 |
| Evropa T 04 | 2* | 7+9 | 2+12 |
Payne i drugi autori sačinili su sistem bodovanja (Glu 1 quality score) za HMW-GS, gdje su individualne subjedinice obilježene brojevima i označavaju ocjene gradirane prema kvalitetu. Ocjene se kreću od 3 do 10, gdje 10 označava najbolji kvalitet. Dobijeni rezultati nisu rangirani na ovaj način, budući da su veoma raznoliki i neujednačeni po sortama, te se pretpostavilo da ne bi dali adekvatnu sliku kvaliteta ispitivanih kultivara.
Kako se kombinacija HMW-GS 5+10 nasuprot 2+12 veže za kvalitet glutena, uobičajeno je da se podjela uzoraka vrši na osnovu ovih kombinacija (Faergestad et al, 2000, Tronsmo et al, 2002, 2003a, 2003b, 2003c, Aamodt et al, 2003; Aamodt et al, 2004).
Na osnovu ove podjele, sorte pšenice koje sadrže 5+10 kombinaciju Glu-D1, Marija, Renesansa i Pobeda bi se svrstale u sorte, odnosno, uzorke sa jakim glutenom, dok bi se uzorci Tina, Sana BL 04, Rusija BL 03 i Evropa, koji sadrže 2+12 kombinaciju Glu-D1, svrstali u grupu sorti, odnosno, uzoraka sa slabim glutenom.
Kombinacija subjedinica 5+10 i 2+12 u jednom kultivaru je rijetka pojava. Ovo se uočava kod sorti Tina, Renesansa, Sana i Rusija. Ista kombinacija alternativnih alela u pojedinim sortama je uočena i opisana u istraživanju F a e r g e s t a d et al (2004).
Ovo može biti posljedica udjela neke druge sorte u ispitivanom materijalu.
Međutim, moguće je da kultivar sadrži različite biotipove, što znači da roditeljske, odnosno, izvorne sorte, kod ukrštanja sadrže 5+10 (jedan roditelj) i 2+12 (drugi roditelj). Kod prve generacije nakon ukrštanja pojava heterozigota je frekventna. Moguće je da se pojavljuju ovakvi biotipovi u slučaju kada se selekcijom proizvode heterozigotne biljke, koje imaju i 5+10 i 2+12 1D kodirane HMW-GS, i koriste se za daljnje razmnožavanje i testiranje u uzgojnoj liniji. U narednoj generaciji biljke homozigoti i za 2+12 i za 5+10 pojavit će se u istoj sorti (Uhlen, 2006).
Tipično za biotipove kao što je ovaj je da biljke vizuelno morfološki izgledaju isto (homozigote), iako sadrže različite alele na lokusima HMW gluteninskih subjedinica. Ovo nije moguće otkriti u uzgojnim programima gdje se ne koristi kontrola sadržaja HMW-GS, elektroforetski. Činjenica je da su takvi biotipovi prilično česti, posebno kod starijih kultivara (U h l e n, 2006).
Karakterizacija HMW-GS pojedinih uzoraka pšenice pokazala je kako se radi o raznolikom materijalu pšeničnog zrna. To je bio razlog ponavljanja ispitivanja na pojedinačnim zrnima (pet zrna) na uzorcima: Marija BL 03, Liberta BL 03, Liberta BL 04, Tina BL 03, Tina T 03, Sana BL 03 i Renesansa BL 03.
Karakterizacija HMW-GS na ovim uzorcima pšenice, na pojedinačnim zrnima, predstavljena je u tabeli 3.
Tabela 3.
HMW GS determinated on individual grain
| Slika/Pozicija Fig./Position | Uzorak Sample | Gtfi-Al | Glu-Bl | Glu-Dl |
| 1/1 | Marija BL 03 | 2* | 7 | 5+10 |
| 1/2 | 1 | 2* | 7 | 5+10 |
| 1/3 | 1 | 2* | 6+8 | 5+12 |
| 1/4 | 1 | 2* | 7 | 5+10 |
| 1/5 | 1 | 0 | 6+8 | 5+12 |
| 1/6 | Liberta BL 03 | 2* | 6+8 | 5+12 |
| 1/7 | 1 | 2* | 6+8 | 5+12 |
| 1/8 | 1 | 0 | 6+8 | 5+12 |
| 1/9 | 1 | 0 | 6+8 | 5+12 |
| 1/10 | 1 | 1 | 7+8 | 2+12 |
| 1/13 | Liberta BL 04 | 2*/0 | 6+8 | 2+12 |
| 1/14 | 1 | 2* | 7 | 5+10 |
| 1/15 | 1 | 2*/0 | 6+8 | 2+12 |
| 2/1 | 1 | 2*/0 | 6+8 | 2+12 |
| 2/2 | 1 | 2*/0 | 6+8 | 2+12 |
| 2/3 | Tina BL 03 | 2* | 6+8 | 5+12 |
| 2/4 | 1 | 1 | 7+8 | 2+12 |
| 2/5 | 1 | 2* | 6+8 | 5+12 |
| 2/6 | 1 | 2* | 6+8 | 5+12 |
| 2/7 | 1 | 1 | 7+8| | 2+12 |
| 2/10 | Tina T 03 | 2* | 7 | 5+10 |
| 2/11 | 1 | 2* | 7 | 5+10 |
| 2/12 | 1 | 2* | 7 | 5+10 |
| 2/13 | 1 | 2* | 7 | 5+10 |
| 2/14 | 1 | 2* | 7 | 5+10 |
| 3/1 | Sana BL 03 | 2* | 6+8 | 5+12 |
| 3/2 | 1 | 2* | 6+8 | 5+12 |
| 3/3 | 1 | 2* | 7 | 5+10 |
| 3/4 | 1 | 2* | 7 | 5+10 |
| 3/5 | 1 | 2* | 6+8 | 5+12 |
| 3/6 | Renasansa BL 03 | 0 | 7 | 5+10 |
| 3/7 | 1 | 2*/0 | 6+8 | 2+12 |
| 3/8 | 1 | 0 | 7 | 5+10 |
| 3/9 | 1 | 2* | 7 | 5+10 |
| 3/10 | 1 | 2*/0 | 6+8 | 2+12 |
Rezultati karakterizacije HMW-GS na pojedinačnim zrnima kod prikazanih uzoraka pšenice pokazali su neujednačen sadržaj HMW-GS. Naročito su interesantni rezultati sa kombinacijama Glu-D1 5+12 u pojedinačnom zrnu, i to na uzorcima sa jedne lokacije i jedne uzgojne godine: Marija, Liberta, Tina i Sana, što svakako predstavlja problem pri razumijevanju i tumačenju ove pojave.
Ova kombinacija mora nastati rekombinacijom između 5+10 i 2+12 subjedinica. Ovakvi slučajevi su već referisani, mada se čini kako je ova pojava prilično rijetka. Međutim, ako se dese, ovakve rekombinacije mogu ostati sačuvane u kultivaru, a takođe se mogu prenositi na nove kultivare (U h l e n, 2006).
Slike gelova sa karakterizacijom HMW-GS na pojedinačnim zrnima prikazane su na slikama 1, 2 i 3.
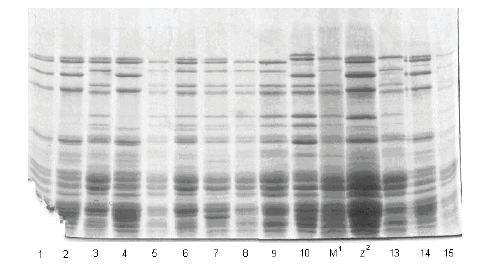
Prikaz HMW-GS na 5 pojedinačnih zrna uzoraka Marija BL 03, Liberta BL 03 i Liberta BL 04
Slika/Fig 1
Gel slabs of HMW GS on 5 individual grains of samples Marija BL 03, Liberta BL 03 i Liberta BL 04
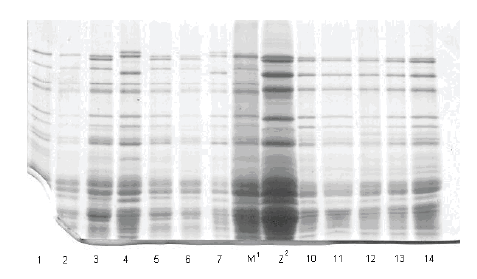
Prikaz HMW-GS na 5 pojedinačnih zrna uzoraka Liberta BL 04, Tina BL 03 i Tina T 03
Slika /Fig. 2
Gel slabs of HMW GS on 5 individual grains of samples Liberta BL 04, Tina BL 03 i Tina T 03
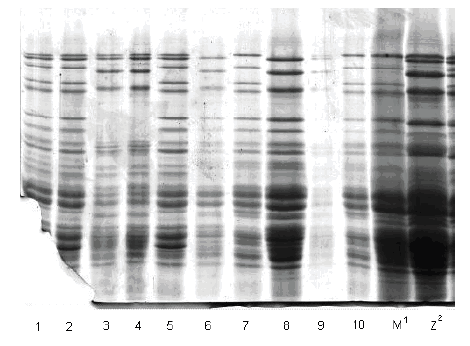
Prikaz HMW-GS na po 5 pojedinačnih zrna uzoraka Sana BL 03, Renesansa BL 03
Slika /Fig. 3
Gel slabs of HMW GS on 5 individual grains of samples Sana BL 03, Renesansa BL 03
Zaključci
Na osnovu provedenog istraživanja koja su obuhvatala karakterizaciju HMW gluteninskih subjedinica na aktualnim sortama pšenice u BiH izvode se slijedeći zaključci:
− Na osnovu karakterizacije HMW gluteninskih subjedinica, sorte pšenice su svrstane u uzorke sa jakim glutenom, odnosno one koje sadrže Glu-D1 5+10 kombinaciju: Marija, Renesansa, Pobeda;
− Sorte pšenice koje sadrže Glu-D1 kombinaciju 2+12, odnosno sorte sa slabim glutenom, su: Tina, Sana i Evropa;
− Kombinacija subjedinica glutenina 2+12 i 5+10 u jednom kultivaru je relativno rijetka i nađena je kod sorti: Tina, Renesansa, Sana i Rusija, što može biti posljedica udjela neke druge sorte u ispitivanom materijalu;
− Karakterizacija HMW gluteninskih subjedinica na pojedinačnim zrnima pokazala je prisustvo Glu-D1 5+12 kombinacije, što je veoma rijetka pojava i u literaturi nedovoljno obrađena. Ova neuobičajena kombinacija nađena je kod uzoraka pšenice Marija, Liberta, Tina i Sana sa lokacije Banja Luka i 2003. uzgojne godine, što svakako predstavlja svojevrstan problem pri razumijevanju ove pojave;
− Karakterizacija HMW gluteninskih subjedinica u aktuelnim sortama pšenice koje se uzgajaju na području BiH odražava nepovoljnu situaciju u sortimentu, te je neophodno uvoditi nove i/ili „čiste“ sorte, koje bi obezbijedile ujednačenu sirovinu, odnosno ujednačen kvalitet, čemu bi doprinijelo uvođenje permanentne kontrole HMW gluteninskih subjedinica tokom samog uzgoja.
Literatura
Aamodt, A., E. M. Magnus, E. M. Faergestad (2003): Effect of flour quality, ascorbic acid, and DATEM on dough rheological parameters and heart loaves characteristic. Journal of Food Science 68(7):2201-2210.
Aamodt, A., E. M. Magnus, E. M. Faergestad (2004): Effect of Protein Quality, Protein Content, Bran Addition, DATEM, Proving Time, and Their Interaction on Heart Bread. Cereal Chemistry. 81(6):722-734.
Eliasson, A. C. (1990): Rheological Properties of Cereal Proteins. 67-110 U: Dough Rheology and Baked Product Texture. (Ed.) Hamed, A.F. and Faubion, J.M. New York, USA.
Faergestad, E. M., E. L. Molteberg, E. M. Magnus (2000): Interrelationships of protein composition, protein level, baking process and characteristic of heart bread and pan bread. Journal of Cereal Science 31:309-320.
Faergestad, E. M., N. S. Fleate, E. M. Magnus, K. Hollung, H. Martens, A. K. Uhlen (2004): Relationships between storage protein composition, protein content, growing season and flour quality of bread wheat. Journal of the Science of Food and Agriculture 84:877-886.
Gupta, R. B., F. Bekes, C. W. Wrigley (1991): Prediction of Physical Dough Properties from Glutenin Subunits Composition in Bread Wheats: Correlation Studies. Cereal Chemistry. 68(4):328-333.
Gupta, R. B., J. G. Paul, G. B. Cornish, G. A. Palmer, F. Bekes, A. J. Rathjen (1994): Allelic Variation at Glutenin Subunit and Gliadin Loci, Glu-1, Glu-3 and Gli-1, of Common Wheats. I. Its Additive and Interaction Effects on Dough properties. Journal of Cereal Science 19:9-17.
Kaluđerski, G., N. Filipović (1998): Metode ispitivanja kvaliteta žita, brašna i gotovih proizvoda, Tehnološki fakultet, Zavod za tehnologiju žita i brašna, Novi Sad.
Lafiandra, D., D. D. Kasarda (1985): One- and Two-Dimensional (Two-pH) Polyacrylamide Gel Electrophoresis in a Single Gel: Separation of Wheat Proteins. Cereal Chemistry. 62(5):314-319.
Lookhart, G. L., M. L. Martin, E. Mosleth, A. K. Uhlen, R. C. Hoseney (1993): Comparison of High-molecular-weight Subunits of Glutenin and Baking Performance of Flours Varying in Bread-making Quality. Lebensm.-Wiss. u.-Technol. (26):301-306.
MacRitchie, F. (1999): Wheat proteins: Characterization and Role u Flour Functionality. Cereal Food World. Vol. 44. NO. 4.
Payne, P. I. (1987): Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annual Review of Plant Physiology. 38:141-153.
Payne, P. I., C. N. Law, E. E. Mudd (1980): Control of homoeologous group 1 chromosomes of the high-molecular-weight subunits of glutenin, a major protein of wheat endosperm. Theoretical and Applied Genetics. 58:113-20.
Payne, P. I., G. J. Lawrence (1983): Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular weight subunits of glutenin in hexaploid wheat. Cereal Research Communications 11 (1) 29-35.
Payne, P. I., K. G. Corfield, L. M. Holt, J. A. Blackman (1981): Correlations Between the Inheritance of Certain High-molecular Weight Subunits of Glutenin and Bread-making Quality in Progenies of Six Crosses of Bread Wheat. Journal of the Science of Food and Agriculture. 32:51-60.
Payne, P. I., L. M. Holt, A. F. Krattiger, J. M. Carrillo (1988): Relationships Between Seed Quality Characteristic and HMW Glutenin Subunits Composition Determined using Wheats Grown in Spain. Journal of Cereal Science. 7:229-235.
Payne, P. I., L. M. Holt, M. G. Jarvis, E. A. Jackson (1985): Two-Dimensional Fractionation of the Endosperm Proteins of Bread Wheat (Triticum aestivum): Biochemical and Genetic Studies. Cereal Chemistry. 62(5):319-326.
Pena, R. J., H. Gonzalez-Santoyo, F. Cervantes (2005): Relationship between Glu-D1/Glu-B3 allelic combinations and bread-making quality-related parameters commonly used in wheat breeding. www.rsc.org/pdf/books/gluten_proteinsamend.pdf.
Shewry, P. R., N. G. Halford, A. S. Tatham (1992): High molecular weight subunits of wheat gluten. Journal of Cereal Science. 15:105-120.
Shewry, P.R., A.S. Tatham, P. Lazzeri (1997): Biotechnology of Wheat Quality, Journal of the Science of Food and Agriculture; 73 397-406.
Tronsmo, K. M., E. M. Faergestad, Å. Longva, J. D. Schofield, E. M. Magnus (2002): A study of how size distribution of gluten proteins, surface properties of gluten and dough mixing properties relate to baking properties of wheat flours. Journal of Cereal Science 35:201-214.
Tronsmo, K. M., E. M. Faergestad, J. D. Schofield, E. M. Magnus, (2003c): Wheat protein quality in relation to baking performance evaluated by the Chorleywood bread process and a heart bread baking test. Journal of Cereal Science 38:205-215.
Tronsmo, K. M., E. M. Magnus, E. M. Faergestad, J. D. Schofield (2003a): Relationships between gluten rheological properties and heart loaf characteristic. Cereal Chemistry 80(5):575-586.
Tronsmo, K. M., E. M. Magnus, P. Baardseth, J. D. Schofield, A. Aamodt, E. M. Faergestad (2003b): Comparison of small and large deformation rheological properties of wheat dough and gluten. Cereal Chemistry 80(5):587-595.
Uhlen, A. K. (2006): Privatna korespodencija.
Uhlen, A. K. (1990): Quantitative analysis of the high molecular weight subunits of glutenin present in Norwegian wheats. Norwegian Journal of Agricultural Science. 4:19-26.
Wrigley, C. A., J. A. Bietz (1988): Proteins and amino acids. Pages 159-275 u Wheat: Chemistry and Technology. Volume I. Y. Pomeranz, Ed. AACC. USA.